ЗАРОДЫШЕВЫЕ ЛИСТКИ
Спасибо нашим инвесторам из казино онлайн
ЗАРОДЫШЕВЫЕ ЛИСТКИ , слои или пласты (нем. Keimblatter, франц. feuillets ger-minatifs, англ. germinal layers), основной эмбриологический термин, которым обозначают слои эмбриональных клеток, образующие в раннем стадии развития тела зародыша и имеющие в большинстве случаев эпителиальный характер.
Принято различать три зародышевых листка:
- наружный (эктодерма, эктобласт, эпибласт, кожно-чувствующий листок);
- внутренний (энтодерма, энтобласт, гипобласт, кишечно-железистый листок);
- средний (мезодерма, мезобласт) (рисунки 1, 2, 3).
Из них первые два образуются раньше, и к ним позднее присоединяется третий. Наружный зародышевый листок состоит обыкновенно из высоких светлых клеток, похожих на цилиндрический эпителий; внутренний зародышевый листок может состоять из крупных клеток, выполненных желточными пластинками и образующих местами компактные массы (амфибии), или, наоборот, из клеток, уплощенных наподобие плоского эпителия (птицы, млекопитающие); средний зародышевый листок в начале образования может состоять из рыхло расположенных веретенообразных или звездчатых клеток, которые впоследствии слагаются в эпителиальный слой. Некоторые авторы считают средний зародышевый листок за два листка (париетальный и висцеральный мезобласт, иначе – кожно-мышечный и кишечно-волокнистый листки), так как на большом протяжении он является расщепленным.
Зародышевые листки позвоночного животного
На рисунках 1, 2 и 3 приведены зародышевые листки следующих позвоночных животных – тритона, цыплёнка и колика:
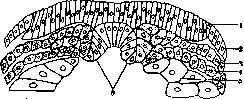
Рисунок 1. Зародышевые листки тритона; 1 – медулярная пластинка; 2 – эктодерма; 3 – париетальный листок мезодермы; 4 – висцеральный листок мезодермы; 5 – энтодерма; 6 – хорда (по Hertwig’у).
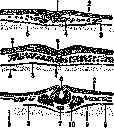
Рисунок 2. Зародыш, листки цыпленка. Разрезы бластодермы трех следующих друг за другом стадии – А, В, С: 1 – первичная бороздка; 2 – эктодерма; 3 – энтодерма; 4 – мезодерма; 5 – желток; 6 – зачаток нервной трубки; 7 – хорда; 8 – полость тела; 9 – мезодерма полости тела; 10 – сомит (по Meisenheimer’у).
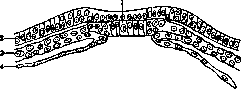
Рисунок 3. Зародышевые листки кролика: 1 – хорда; 2 – эктодерма; 3 – мезодерма; 4 – энтодерма (по Beneden’у).
Учение о зародышевых листках, их возникновении и дальнейшей судьбе проходит через всю историю эмбриологии; после Дарвина оно тесно связывается с эволюционным учением и становится основой сравнительной эмбриологии; в начале 80-х годов братья Гертвиги (Hertwig) приводят его в стройную систему, в каком виде оно обыкновенно и излагается в учебниках. Но с другой стороны оно подвергается сильной критике, и в настоящее время взгляды на зародышевые листки далеко не приведены к единству. Поэтому надлежащее представление о зародышевых листках без знакомства с историей вопроса составить трудно.
Исторические данные
К. Фр. Вольф (К. Fr. Wolff), заложивший своими исследованиями над развитием цыпленка основу современной эмбриологии, описал (в 1768 году) развитие кишечного канала из зачатка, имеющего вид кожицы или листка, который затем свертывается в трубку, и высказал предположение, что по тому же типу развиваются и прочие системы зародыша; нервная, мышечная, сосудистая. Через 50 лет Пандер (Pander; 1817 г.), исследуя бластодерму цыпленка на 12-м часу насиживания, описал в ней два тонких слоя: серозный и слизистый листки; между ними впоследствии развивается третий – сосудистый. По следам Пандера пошел К. Э. Бэр (1828 – 1837 гг.), который нашел, что два первичных листка (анимальный и вегетативный) в дальнейшем расщепляются каждый на два: из наружного, анимального, образуются кожный и мускульный листки, из вегетативного – сосудистый и слизистый. Впоследствии они свертываются в трубки, образуя первичные органы. Дальнейшие исследования над зародышевом листком цыпленка принадлежат Ремаку (Remak; 1851 г.), который различал всего три листка, называя их по физиологическому значению: наружный – чувствующим, внутренний – трофическим и средний – моторно-герминативным. Средний листок расщепляется на два только по бокам (боковые пластинки); он образует кожно-волокнистый и кишечно-волокнистый листки, ограничивающие полость тела. В то же время зоологи Гексли (Huxley; 1849 г.) и Олмен (Allman; 1853 г.) указали на гомологию между двумя первыми зародышевыми листками и слоями тела у низших беспозвоночных (кишечно-полостных); Олмену принадлежат термины «эктодерма» и «энтодерма», получившие широкое распространение и вытеснившие термины прежних эмбриологов. Обширные исследования над развитием разных классов беспозвоночных и ланцетника были произведены русским ученым А. Ковалевским; они доставили фактический материал для теорий Рей Ланкестера (Ray Lankeater; 1873 г.) и Геккеля (Haeckel; 1874 г.), связавших эмбриологию с филогенией. Эти ученые предполагали, что простейшая форма, давшая в процессе эволюции начало всем прочим беспозвоночным и позвоночным, состояла из двух слоев, которые затем проявляются в течение развития всех животных в виде двух первичных листков. Рей Ланкестер считал такой формой планулу – бластулу, у которой от клеточного слоя отщепляется внутрь второй листок; вследствие прорыва стенки полость планулы сообщается с внешней средой и превращается в первичную кишку. Геккель видел первичную форму в гаструле, образовавшейся путем впячивания, и назвал ее «гастреа» (Gastraeatheorie). Переход двуслойной формы в трехслойную совершается путем отщепления клеток от обоих листков. Теория Геккеля получила широкое распространение, при чем эмбриологи направляли усилия на то, чтобы доказать возникновение двух первых листков путем процесса впячивания (в первых изданиях «Lehrbuch der Entwicklimgsgeschichte» О. Гертвига этот способ образования последовательно проводится для всех позвоночных).
Дальнейшие работы были направлены на изучение среднего зародышевого листка, который вследствие своей разнородности представлял трудности для понимания; они были преодолены трудами Оскара и Рихарда Гертвигов (1881 г.), создавших теорию целома (Coelomtheorie), аналогичную теории гастреи. Бр. Гертвиги прежде всего исключили из состава среднего зародышевого листка мезенхиму (клеточные группы, выделяющиеся из обоих листков и дающие начало соединительной ткани и крови), оставив название мезодермы только за участками, носящими эпителиальный характер, а затем поставили в связь образование мезодермы с развитием полости тела (целом). За образец было взято развитие ланцетника (Amphioxus), изученное Ковалевским и Гачеком (Hat schek), где эта связь выступает с полной ясностью (рисунок 4).
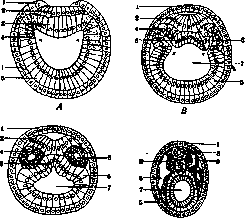
Рисунок 4. Образование мезодермы у ланцетника (А, В, С и D): 1 – эктодерма; 2 – медулярная пластинка; 3 – хорда: 4 – мезодерма; 5 – энтодерма; 6 – полость тела; 7 – полость кишечника; 8 – нервная трубка; 9 – сомит; 10 – место впячивания полости тела (по Hatschek’у).
На известной стадии первичная энтодерма гаструлы дает ряд мешковидных выпячиваний по обе стороны от средней оси – это зачатки полости тела, выстланные мезодермой. В дальнейшем они углубляются между эктодермой и энтодермой и разделяются на участки: проксимальные образуют сомиты (первичные позвонки), дистальные сливаются с последующими и предыдущими, образуя полость тела, расположенную между листками мезодермы – париетальным и висцеральным. Таковы ближайшие производные мезодермы. Этот же способ образования наблюдается у тритона (рисунок 5); у других он затемняется, так как мезодерма вырастает в виде сплошных масс, впоследствии расщепляющихся. Дело еще более осложняется тем, что у селахий, рептилий и птиц мезодерма развивается из двух – мезо и эктодермы (рисунок 2), но, если рассматривать первичную полоску птиц как бластопор и обращать внимание на углубление в узелке Гензена, можно образование мезодермы и здесь связать рядом постепенных переходов с основной схемой.
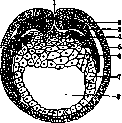
Рисунок 5. Образование мест (периферическая мезодермы у тритона: 1 – бластопор; 2 – пари – и аксиальная, при чем париетальный листок в области первичной дермы;. 3 – желточная полоска вырастает от пробки; 4 – висцеральный листок мезодермы; 5 – эктодерма; 6 – желточные клетки; 7 – энтодерма; 8 – полость кишечника (по Hertwig’у).
Учение о зародышевых листках на основе теории гастреи, целома и бластопора (Urmundtheo-rie) в законченном и стройном виде излагалось в упомянутом учебнике О. Гертвига, который представляет собой лучший памятник сравнительной эмбриологии позвоночных того периода, когда идеи эволюции начали завоевывать признание широких масс естествоиспытателей, не утративший своего значения и в настоящее время. Критика учения о зародышевых листках, не имевшая особого успеха в XIX в., в настоящее время привлекает к себе больше внимания в связи с изменением курса эмбриологии, перешедшей от описания и сравнения к выяснению причин развития при помощи эксперимента. Основное возражение против учения о зародышевых листках было дано еще Рейхертом (Reichert; 1843 г.), который вместо листков выдвигал на первый план зачатки органов (первичные органы), возникающие или прямо как таковые или по нескольку вместе в общем зачатке. В противоположность зародышевым листкам эти первичные органы не являются строго фиксированными понятиями и у разных животных разнятся в числе, форме и положении. В последующее время главные удары критики были обращены на средний зародышевый листок (Kleinenberg, 1886 г.; Bergh, 1896 г.), который и у позвоночных, а в особенности у беспозвоночных, нередко представляет совокупность совершенно разнородных зачатков и как единый листок не существует. Расчленение мезенхимы и мезодермы равным образом не может быть проведено во всем животном царстве и наталкивается на многочисленные противоречия. Главным противником учения о зародышевых листках в последнее время является зоолог Мейзенгеймер (Meisenheimer), всецело разделяющий точку зрения Рейхерта. Но, признавая полную основательность возражений против среднего зародышевого листка, вряд ли можно согласиться с вычеркиванием самого термина «зародышевые листки», так как эктодерма и энтодерма существуют как вполне определенные морфологические образования и бросаются в глаза каждому, изучающему развитие. Иное дело их образование: они могут возникать и действительно возникают у разных животных различным образом в зависимости от количества желтка и других причин, поэтому поддерживать в полной мере теорию Гертвигов не представляется возможным.
Судьба зародышевых листков и их специфичность
Уже первыми исследователями было выяснено в общих чертах, каким органам или частям их дает начало каждый зародышевый листок, иначе говоря, их «проспективное значение».
Развитие органов из трёх зародышевых листков:
- Наружный зародышевый листок производит нервную систему, эпидермис кожи, эпителий и гладкие мышцы кожных желез, эпителий слухового органа, носовой полости, переднего отдела полости рта (включая железистую часть мозгового придатка и эмаль зубов), анальной части прямой кишки, хрусталик, эпителий амниона.
- Внутренний зародышевый листок производит эпителиальную выстилку кишечного канала и образующиеся в нем железы, включая печень и поджелудочную железу.
- Средний зародышевый листок, собственно мезодерма, в области сомитов дает мускулатуру тела (миотом) и соединительную ткань (склеротом), в области нефротома – выделительные органы; мезодерма, выстилающая полость тела, образует его эндотелий (мезотелий) и эпителиальные части половых желез.
Первичные половые клетки в некоторых случаях могут помещаться в энтодерме и оттуда передвигаться в половой валик. Что касается мезенхимы, то она образует клеточные элементы соединительной ткани и кровь, хотя первые зачатки крови некоторые авторы производят из энтодермы. В разграничении мезодермы и мезенхимы полной ясности не существует. Учение о судьбе зародышевых листков было впоследствии дополнено положением об их гистологической специфичности, согласно которому эктодерма, энтодерма, мезодерма и мезенхима обладают ограниченной «проспективной потенцией» и могут производить только определенные виды клеток и тканей. Например, эктодермальный эпителий никогда не может дать начало соединительной ткани или эпителии энтодермальных желез – лейкоцитам. Противоречащие этому утверждения Реттерера (Retterer) о переходе эпителия крипт в лейкоциты или Штёра (Stohr) о возникновении лимфоцитов кожной железы из эпителиального зачатка, рассматривались с недоверием и заставляли предполагать ошибки в наблюдении. На этом же основании в последнее время пытаются проводить разницу между эндотелием сосудов и брюшины: первый как производное мезенхимы может дать начало элементам крови, тогда как мезодермальный эпителий брюшины (мезотелий) к этому не способен (Максимов). Хотя доказанное происхождение гладких мышц желез от экто-дермального и энтодермального эпителия и пробивало брешь в учении о строгой специфичности листковых дериватов, но в общем оно продолжает господствовать и поныне.
Рисунок 6. Продольный разрез зародыша: 1 – Trito cristatus в области сомитов; 2 – сомиты образованы из эктодермы Trilo назначались гистологами alpestris (по Mangold’у).
Вопрос о судьбе зародышевых листков на ранних стадиях развития решается в новейшее время путем эксперимента. Шпеман и Мангольд (Mangold), трансплянтируя различные участки от зародышей пигментированных тритонов (Trito taeniatus) лишенным пигмента (Trito cristatus) (что давало возможность проследить их судьбу), нашли, что в стадии бластулы участки анимального, вегетативного полюсов и промежуточной зоны детерминированы, то есть дают начало определенным листкам, но в стадии гаструлы сформировавшиеся листки не обладают специфичностью. Трансплантированные участки эктодермы могли входить в состав кишечника или наряду с мезодермой давать начало сомитам (рисунок 6). Отсюда делают вывод, что зародышевые листки, не обладая специфичностью, имеют значение только как топографические понятия. В то же время в поздних стадиях гаструлы намечающиеся зачатки органов являются уже детерминированными, и участок мозговой пластинки например везде производит мозг. Экспериментальное изучение гистологической специфичности в прижизненных культурах тканей в общем приводит к тем же результатам.
Литература: Гертвиг О., Элементы эмбриологии, Харьков, 1928; Corning H., Lehrbuch der Kntwicklungsgescbichte des Menschen, Munchen – Wiesbaden, 1921; Mangold 0., Die Bedeutung der Keimblatter in der EntwicMung, Naturwissen-schaften, Band XIII, 1925; Meisenheimer J., Entwicklungsgeschichte der Tiere, Lpz., 1908; он же, Ontogenie (Handworterbuch d. Naturwissenschalten, B. VII, Jena, 1912). В. Карпов.