СЫПНОЙ ТИФ
Спасибо нашим инвесторам из казино онлайн
СЫПНОЙ ТИФ
. Содержание: Экспериментальный С. т.& 182 Сыпнотифозный вирус.&185 Пути и способы распространения С. т.&188 Специфическая профилактика и серотерапия Ст.& 192 Статистика и географическое распространение Ст.&195 Патологическая анатомия С. т.&201 Клиника ». т.&210 Борьба с С. т.&223 Инфекции, сходные с С. т.&226 Сыпной тиф (typhus exanthematicus) (от греч. typhos—дым, туман, оглушение), острое, передающееся вшами инфекционное заболевание, высоко заразительное, циклическое, при котором характерными симптомами являются: быстрое начало, макулезная сыпь, глубокое поражение центральной нервной и периферической кровеносной системы, постоянная лихорадка, разрешающаяся между второй и третьей неделей укороченным лизисом или критически. Синонимы: голодный, тюремный, военный тиф. История. Первое описание характерных признаков С. т. принадлежит Фракастору (Fracastorius, 1483—1553). Далее следуют труды Мосса, Мантууса, Иордануса, Амбруаза Паре (Moss, Mantuus, Jordanus, Ambroise Pare) и др., где указываются отличительные клин. и эпидемиологические признаки С. т. Однако до средины 19 в. многие врачи, в том числе Вирхов, отрицали существование С. т. как самостоятельной болезни. Вопрос был окончательно разрешен благодаря тщательным исследованиям Дженнера (Jemier, 1850), Гризин-гера (Gricsinger, 1856), Мерчисона, Вундерли-ха, Куршмапа (Murchison, «Wunderlich, Cursch-гаапп) и др., доказавших анализом клиники и эпидемиологии сыпного тифа, что он представляет особое специфическое заболевание, ясно отличающееся от других тифов: брюшного и возвратного.
Экспериментальный сыпной тиф.
Мочутковский в 1900 г., удачно заразив себя кровью сыпнотифозного б-ного, представил доказательства того, что инфекционное начало при С. т. находится в крови. Опыт Мочутков-ского был повторен и подтвержден Отеро, Иерсеном, Вассалем (Otero, Yersin, Vassal) и др. В 1909 г. Шарль Николь, Конт и Консей (Ch. Nicolle, Cornpte, Conscil) в Тунисе воспроизвели экспериментальный С. т. у обезьян, заражая их кровью сыпнотифозных б-ных. Такие же результаты были в 1909 г. получены Андерсоном и Гольдбергером (Anderson,. Gold-berger) в Мексике. В 1910—-11 гг. Гавиньо и Жирар (Gavino, Girard) установили, что мор – ские свинки подобно обезьянам могут быть экспериментально заражены сыпнотифозной кровью и дают характерную картину заболевания. Инфекция у свинок, как и у обезьян, способна поддерживаться неограниченно долгое время пассажами на свежих животных. Наблюдения Гавиньо и Жирара были вскоре подтверждены Шарлем Никол ем с сотрудниками, развиты и углублены последующими работами многочисленных авторов.—Т. о. в руках исследователей Ст. оказался ценный способ изучения природы этой инфекции на легко доступном материале. Техника воспроизведения экспериментального С. т. у обезьян и свинок
1 3 5 7 ‘J II 13 15
1? ]9 21 23 25
тшт
щ
т
t-
.1″-^
y-iv
1 3 5
1
9 11 13 15 17 19 21 23 25 ■ i i «; » 40;Q
\Ч \
‘
!._Xl-
1 -\
39;°
\ N \
■
«I
rW ч
IT
jo
Ww^~~vr^W
1 . – . If – JJ
S7J5 1 ■ 1 1
нотифозного. материала, к-рый вводится в виду его ненадежной чистоты не в брюхо, а под кожу животным. Экспериментально С. т. способен неограниченно долго пассироваться на новых животных. Материалом для пассажей служат кровь, мозг или селезенка, взятые в разгар лихорадки. Обычно для пассажей животное убивается не позже 3—4-го дня лихорадки, стерильно добытый мозг (resp. селезенка и другие органы) растирается в 0,85%-ном NaCl и впрыскивается в брюхо свежим животным. При пассажах вирус адаптируется к данному животному виду, инкубация укорачивается, лихорадка становится более длительной и высокой, процент животных, восприимчивых к пассажному вирусу, резко нарастает, достигая 90% и выше для обезьян и свинок. Из внутренних органов пассажных животных особенно богат вирусом большой головной мозг (серое вещество). У некоторых сыпнотифозных щтам-
II 13 15 17 1(|
21
2325
УЧ;
ЧК_:
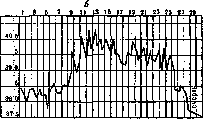
мов уже 7и рис 1 Различные Формы сыпного тифа у морских свинок: с^-бессимптом-пая – б—легкая; в—типическая; г—тяжелая [толстая линия—первичное заражение, тонкая—вторичное (через i'/is мес.)]. состоит в введении им подкожно или внутри – давая уже на 3 брюшинно 2—4
см3
крови сыпнотифозного больного, взятой стерильно из локтевой вены в разгаре заболевания. Инкубация в среднем равна 8—12 дням (колебания от 3 до 31 дня), период лихорадки длится около 1—I1/» недель (колебания от 2 до 16 дней). Лихорадка преимущественно постоянного типа. Она разрешается укороченным лизисом, причем t° иногда падает ниже нормы и приходит к норме только через 1—Здня. Никаких иных симптомов заболевания кроме лихорадки и легкого падения в весе ни у свинок ни у обезьян не наблюдается. Экспериментальный С. т. у названных животных может быть тяжелым, средним или легким, как это видно на прилагаемых температурных кривых (рисунок 1). Наконец в нек-рых случаях экспериментальная инфекция может протекать совсем бессимптомно в виде безлихорадочного сепсиса, как это установлено в 1919 г. Ш. Николем и Лебайи (Lebailly) («infection inapparente»). Кровь таких животных, привитая свежим, дает у них типическое лихорадочное заболевание. Обезьяны и свинки, заражаемые кровью сыпнотифозных больных, не все оказываются восприимчивыми к инфекции человеческим вирусом. Невосприимчивость • у обезьян наблюдается иногда в 22% ел., у свинок—в 44% (Андерсон и Гольдбергер). Цифры эти имеют относительное значение и зависят от происхождения и индивидуальности сыпнотифозного вируса (Барыкин и сотрудники). Экспериментальное заражение обезьян и свинок возможно не только сыпнотифозной кровью от больных людей, но и органами (особенно мозгом и селезенкой) от трупного сын – головного мозга пассажной свинки содержит инфекционную дозу вируса для свежих свинок. Другие штаммы менее вирулентны, особенно штаммы, выделенные от людей при G. т. спорадическом или полученные во внеэпидемические сезоны. Такие штаммы при сериальном прохождении через организм свинок не только не усиливаются, а постепенно падают в своей активности, -5-м пассаже бессимптомную экспериментальную инфекцию или же полностью утрачивая способность заражать свинок. В зависимости от индивидуальности пассируемого сыпнотифозного штамма смертность среди пассажных животных колеблется, достигая для одних штаммов 18.4% (штамм Otto и Papamarku для свинок). Исследованиями последнего времени установлено, что к сыпнотифозному вирусу чувствительны суслики (Sper-mophilus), у которых экспериментальная инфекция протекает так же, как у свинок (Lepine, Грюнфельд, Серебряная и Нейман). Изучение экспериментального С. т. у обезьян и свинок показывает, что они гораздо менее восприимчивы к этой инфекции, чем люди. Сыпнотифозная инфекция у них воспроизводится только искусственно в лабораторной обстановке, имеет доброкачественное течение, сопровождается почти единственным симптомом— лихорадкой, дает ничтожную смертность. Еще легче экспериментальный С. т. протекает у кроликов, крыс и мышей. У этих животных, как показывают наблюдения Ш. Николя, Кон-та и Консея’—для кроликов, Дерра (Doer), Отто и Винклера (Winkler)—для крыс, III. Николя—для мышей, лабораторное заражение сыпнотифозным вирусом выражается в бессимптомной форме инфекции. Кровь этих животных, взятая на 7—15-й день после их лабораторного заражения, при введении ее в брюхо свинкам вызывает у последних типический лихорадочный приступ экспериментального С. т. Кролики при заражении их сыпнотифозным мозгом дают положительную реакцию Вейль-Феликса. Попытки заразить С. т. овец, коз,

Рнс» I— А. Гранулемы ниыошнщ мозг} ^ипниткфо. шш! сншшн. Рис ]. Мал« увеличение, ГрвяудематЕй кадоляров и меткого сосуда in центре) Рнс, _. Среднее рнеякченяе. Граблена. гроиол и кроеогалшише в кашпярс, рассеянный глиоаяыя бугорок. Рнс, Д. Среднее ушмпчснис. Гнпнчсская гранулема if л месте кишмяпл. Ри*. I. Средней ушмчеяне. .^фта оокрлт мелкого eeeyjWL Ряе, 5-—W. Rtekelltji ['ruwn/cki
и органа* ЯКДвА, ПОГИ1ПНХ от ci-imioro пира. RkkettsJi ProwflzeJcl лежат Ешугрнклеточно.
акр
.ены и кроений пнет. Рисунок Л. .Члэок из" галопного
мозга.
Рисунок Ei. Клетка: p)csii* спогЮШсиа распялаетея, нпйитл KickeUsia ProwpTCkl. расположенными кяк бы и кисте. Рис, Г. Малок вз селезенки. HJckeU*ld I'rOftd/ekt рассеяна н прогпн.1вэчс «леток, Рнс. ,4. МайОК из pftxua cliorloidcn^ Внутри-и внеклеточное распело - тине Rfeketlala t'mwazckl, Рнс. !). Мазок на цолгя, Вокруг аядонписточнон групп и HlckeUsJa Pre tvatsekt—лросветдздн иротавлаажи. К ст.
ОлпноЛ тиф.
ослов, собак и кур окончились неудачей. Селивановой в 1924—25 гг. удалось воспроизвести экспериментальный С. т. у кур, уток, голубой и цесарок. Подтверждения этих данных до сих пор не последовало. Что касается патологической анатомии экспериментального С. т., то она тщательно изучена рядом авторов: Леви, Отто и Дитрихом, Рицем Бауером, Барыкиным с сотрудниками, Цееленом, Давыдовским (Levy, Dietrich, Ritz, Bauer, Ceelen) и мн. другими. Она по существу ничем не отличается от пат. анатомии человеческого Ст. 'Макроскоп, изменений, характерных для С. т., при экспериментальной инфекции не наблюдается. Микроскопически дело идет об универсальном гранулематозе мелких сосудов и капиляров, особенно центральной нервной системы (головной мозг, Аммопов рог) (см. отдельную таблицу, рис. 1—4). Гранулематоз обнаруживается с 4—5-го дня лихорадки. Инволюция гранулем заканчивается на 2—3-й неделе апире-ксии. Основным отличительным признаком экспериментального сыпнотифозного гранулема-тоза от такого же при естественной инфекции у людей нужно считать преобладание при первом пролиферативных явлений над деструктивными. Тяжесть и распространенность экспериментального сыпнотифозного гранулсматоза зависят: 1) от индивидуальности сыпнотифозного штамма, которым воспроизводится инфекция, 2) от дозы заразного материала и 3) от веса и индивидуальности заражаемого животного. По Дерру и Киршнеру (Kirschner), у свинок, заведомо больных экспериментальным С. т., гранул ематоз не обнаруживается в 15—20% случаев, по Барыкину, Компапеоц, Захарову и Барыкиной—в 33%. При легких и стертых формах экспериментального сыпного типа гранулемы встречаются крайне редко или вовсе отсутствуют. Сыпнотифозный вирус. Экспериментальное воспроизведение С. т. позволило исследователям изучить природу и свойства сыпнотифозного вируса. Так, было установлено, что вирус этот не относится к категории фильтрующихся, хотя и стоит на границе с последними, проходя в нек-рых редких случаях свечи Шамберлана L2 и Берке-фельда V и N. Далее было доказано, что сыпнотифозный вирус крайне нестоек. Он в сыпнотифозной крови погибает при подогревании до 55° в течение 15 минут (Андерсон и Гольд-бергер, da Rocha-IMma), при комнатной t°— через 2—5 дней (Olitzky), при замораживании—через 2 дня (Hamdi), по нек-рым же авторам через 15 дней (Гольдбергер и Андерсон). Глицерин 30—80%-ный, сапонин, карболовая к-та5%-ная, сулема 1°/00-ная быстро убивают сыпнотифозный вирус. Попытки микроскопически обнаружить этот вирус или получить его культуру в течение десятков лет оказывались бесплодными. Ряд исследователей, начиная с Галлера (Haller, 1868) до Анигштейна и Ам-зеля (Anigstein, Amzel, 1927), описывали в качестве возбудителей С. т. самых разнообразных микробов: Protozoa, кокков, палочек, спирохет и пр. Многие из этих микробов выделялись из сыпнотифозной крови и давали с ней реакции иммунитета, но ни один из них при последующей проверке не оказался истинным возбудителем С. т. Такое разнообразие находок при С. т. объясняется той легкостью, с которой сыпнотифозная кровь вследствие множественного гранулематоза, разрушающего мелкие сосуды, наводняется посторонними микробами (Барыкин). Поиски возбудителя С. т. стали на более прочную почву, когда было доказано, что передатчиком Ст. является платяная вошь (Ш. Николь, 1909). Уже в 1910 г. Риккетс и Вильдер (Ricketts, Wilder) обнаружили в кишечнике платяных вшей, заражен-ных мексиканским С. т., овальные и палочковидные образования, напоминающие бактерий геморагической септицемии. Тождественные образования были найдены у сыпнотифозной вши Провацеком (Prowazek; Сербия, 1913), Сер-жаном, Фолеем и Виалаттом (Sergent, Foley, Vialatte; Тунис, 1914). В конце 1914 г. да Ро-ха-Лима, систематически изучая под микроскопом платяных сыпнотифозных вшей, собранных с русских военнопленных в лагере Котт-бусе, установил, что образования эти встречаются в огромных количествах в кишечном эпителии вшей и являются постоянным характерным признаком зараженности последних сыпнотифозным вирусом. Роха-Лима назвал их «Rickettsia Prowazeki» в честь двух вышеупомянутых ученых, видевших их и погибших о
т С. т. при своих исследованиях. Согласно описанию Роха-Лима Rickettsia Prowazeki имеют основную эллиптическую форму. Очень короткие, почти круглые они растут, вытягиваются в длину и затем делятся на два новых элемента. Периоду полного разделения предшествует образование перетяжки, нек-рое •время еще соединяющей молодые особи и придающей всему микробу вид бисквита или гимнастической гири. Наряду с такими формами встречаются палочковидные, окрашивающиеся полюсно, и длинные, вытянутые экземпляры. В расположении на препаратах нет ничего характерного. Отдельные образования иногда располагаются в виде коротких цепочек. Характерно огромное количество образований, напоминающих на мазках чистую культуру микробов. Rickettsia Prowazeki по величине стоят на границе разрешающей способности современных микроскопов. Молодые формы имеют в длину около 0,3—0,4
ft,
делящиеся— около 0,6—0,9
ft.
Их морфология видна на микрофотограмме [см. отдельную таблицу (ст. 199—200), рис. 1 и 2]. Rickettsia Prowazeki окрашиваются с трудом и бледнее банальных микробов. Наиболее надежным для них является метод Гимза, при к-ром они окрашиваются в бледнорозовый с малиновым оттенком цвет (см. отдельную таблицу, рисунки 5—9). По Граму не красятся, самостоятельным движением не обладают, взвешенные в 0,85%-ном растворе хлористого натрия обнарузкивают ясное Броуновское движение. Капсула у них не найдена, хотя нередко на окрашенных по Гимза препаратах они кажутся окруженными прозрачной зоной. Важнейшим отличительным признаком является локализация их в эпителии средней кишки (желудка) вши. Внедряясь в этот эпителий, они энергично, размножаются, растягивают, а потом и разрывают эпителиальную клетку, подвергающуюся глубоким дегенеративным изменениям. Вместе с обрывками клеток Rickettsia Prowazeki попадают в просвет кишечника и выводятся с фекалиями наружу, как это видно на прилагаемой микрофотограмме. Сикора нашла Rickettsia Prowazeki в слюнных железах зарая-генньгх вшей, 187 СЫПНОЙ ТИФ & 188 что оспаривается другими исследователями, Аркрайт и Бекот (Arkwright, Bacot) — в яичниках и яйцах самок вшей. Наиболее убедительные картины сыпнотифозного риккетсиоза у вшей дают сериальные разрезы из их кишечника, приготовленные но способу Сикоры или Вейгля (Sikora, Weigl). Rickettsia Prowazeki найдены в крови и органах сыпнотифозных людей и свинок. Так, Риккетс и Вильдер видели их в крови больных людей, Бессау (Bessau)—в кровяной плазме, Кучинский, Яффе (Kuczynski, Jaffo), Барыкин и Афанасьева—в эндотелии капиляров ■внутренних органов, Вольбах, Тодд и Палф-рей (Wolbach, Todd, Palfrey)—в эндотелии кожных гранулем и т. д. Характерны внутриклеточное расположение Rickettsia Prowazeki и скопления их в виде целых групп [см. отдельн. табл. (ст. 183—184), рис. 5—9]. Вудкок (Woodcock) рассматривает Rickettsia Prowazeki как неживые образования типа гранул, митохондрий или продуктов клеточного распада. Против такого истолкования говорит вся совокупность фактов, доказывающих живую природу Rickettsia Prowazeki и их тесное отношение к этиологии С. т. Rickettsia Prowazeki в отличие от гранул и клеточного распада морфологически и тинкториально единообразны. Они не разрушаются даже крепкой уксусной кислотой, кипящей дест. водой, 2%-ным раствором NaCl, эфиром, ацетоном, толуолом, ксилолом, бензином и спиртом (Эп-штейн). Их переваривает панкреатин. Они являются полноценными антигенами. В пользу их этиологического значения для С. т. может быть представлен ряд веских и хорошо проверенных фактов: 1) Rickettsia Prowazeki, локализованные в кишечном эпителии, встречаются только у вшей, снятых с сыпнотифозных б-ных или накормленных сыпнотифозной кровью; 2) только вши, содержащие Rickettsia Prowazeki, вызывают экспериментальный С. т. с иммунитетом к последующему заражению сыпнотифозной кровью; 3) Rickettsia Prowazeki но своей величине, отношению к t° и другим агентам ведут себя вполне тождественно с вирусом С. т.; 4) Rickettsia Prowazeki дают все реакции иммунитета с сыпнотифозными сыворотками людей и экспериментальных животных; 5) optimum развития Rickettsia Prowazeki, как и вируса С. т., в теле платяной вши лежит около + 32° (Роха-Лима); при t°+23° во вши прекращается развитие как Rickettsia Prowazeki, так и вируса С. тифа. На основании указанных фактов огромное большинство исследователей считает Rickettsia Prowazeki возбудителем С. т. В отличие от банальных микробов Rickettsia Prowazeki не культивируется на обычных лабораторных средах. В 1923 г. Кучинскому, а также Крон-товскому и Гаху удалось поддержать в одной генерации вирус С. т. в эксплантатах селезенки сыпнотифозной свинки по методу Карреля (Carrel). Дальнейшее усовершенствование этой методики применительно к С. т. в руках Сато Кийоши, Нигг и Ландштейн
ера (Sato Kiyoshi, Nigg, Landsteiner), как и др. авторов, привело к окончательному разрешению задачи культивирования сыпнотифозного вируса в тканевых эксплянтатах. Методика авторов обеспечивает получение сыпнотифозной культуры в неограниченном количестве пересевов. Культура сохраняет полностью все свои вирулентные, антигенные и иммуннологические свойства. Мор – фологически она представляет собой культуру Rickettsia Prowazeki. размножающихся в клетках эксплянтата. В некоторых случаях обнаружение этих образований в тканевых культурах требует специальной обработки (Сато Кийоши) или их совсем не удается обнаружить. Тканевые культуры однако не могут дать больших количеств сыпнотифозного вируса и следовательно не могут служить материалом для опытов массового приготовления противо-сыпнотифозной вакцины или сыворотки. Оригинальная методика массового культивирования сыпнотифозного вируса была недавно предложена Зильбером и Доссер, выращивающими этот вирус в симбиозе с другими микробами, в частности с кефирными дрожжами. Авторы получили 16 генераций сыпнотифозного вируса, но биология его повидимому несколько страдает, что выражается утратой им своих антигенных свойств (Барыкин). Интересно отметить, что в дрожжевых культурах Rickettsia Prowazeki не обнаруживаются. Со времени установления значения этих образований в этиологии С. т. морфологически тождественные образования были найдены у вшей, собранных как со здоровых людей (Brumpt), так и с людей, больных волынской лихорадкой (Jimgmann, Кучинский) и рядом других инфекций. В наст, время доказано, что риккетсиозы широко распространены среди сосущих и несосущих кровь насекомых. Они встречаются как у Insecta, так и у Arachnoidea. Однако все эти виды риккетсиозов («Rickett-sienarten») по совокупности признаков, особенно по отношению к кишечному эпителию насекомого, легко могут быть отличимы от Rickettsia Prowazeki (Роха-Лима). Этиологическое значение для С. т. Rickettsia Prowazeki оспаривалось некоторыми авто-’рами. Однако попытка Вейля и Феликса, Фридбергера (Friedberger), Кучинского, Фейги-ной (Fejgin) и других связать этиологию С. т. с культурой В. protcus X19, специфически агдютипирующейся сыпнотифозными сыворотками, оказались несостоятельными. Против этиологического значения этой культуры для С. т. говорят: 1) крайне редкое выделение ее из сыпнотифозной крови, 2) неспособность В. proteus Xle и его вариантов вызывать экспериментальный Ст., 3)отсутствие ее у сыпнотифозных вшей и пр. Самая способность Х-про-теев входить в специфическую аглютинацию с сыпнотифозными сыворотками рассматривается большинством авторов как свойство пара-штамма, приобретаемое ИМ’В результате его сожительства с вирусом, и квалифицируется как парааглютинация. Таким образом, резюмируя данные по этиологии Ст., нужно заключить, что возбудителем этой инфекции являются Rickettsia Prowazeki, хотя в нек-рых исключительных случаях их и не удается обнаружить, что может быть зависит от способности их давать при известных условиях фильтрующиеся невидимые формы.
Пути
и
способы распространения сыпного тифа»
Пути и способы распространения С. т. предопределяются тем основным и бесспорным положением, что передатчиком Ст. является платяная вошь (Pediculus vestimenti). Это впервые было экспериментально установлено на обезьянах в 1909 г. Ш. Николем, Контом и Консеем, а в дальнейшем нашло полное подтверждение как у экспериментаторов, так и у клиницистов и эпидемиологов всех стран света. В пользу главной и решающей роли вши в распространении С. т. могут быть приведены многочисленные доказательства. Из них важнейшими являются следующие: 1) обесвши-вленный тифозный б-ной перестает быть заразительным для окружающих, 2) уничтожение вшей среди населения влечет за собой немедленное исчезновение С. т., 3) вспышки С. т. обычно связаны с холодными временами года, когда население живет более скученно, реже имеет возможность мыться, носит шерстяную и меховую одежду, трудно доступную обес-вшивливанию, и когда благодаря всем указанным сезонным и бытовым условиям вшивость может достигнуть наибольших размеров. Главным передатчиком С. т. является платяная вошь, живущая в нательном белье и переходящая на человека 3—4 раза в день для того, чтобы кормиться его кровью. Рядом с платяной вошью несравненно менее видную, но все же бесспорную роль в распространении С. т. играет головная вошь (Pediculus capitis) и возможно п л о щ и ц а (Pediculus pubis). Передача сыпнотифозного вируса вшами может осуществляться тремя способами: 1) при помощи укусов зараженной вошью здорового человека (Ш. Николь с сотрудниками); 2) при помощи раздавливания эт